7-1トリテルペン



人類にとって重要な穀類、例えばコムギ、トウモロコシ、オオムギ、エンバク、イネなど人類の主食のほとんどは単子葉植物ですが、単子葉植物類ではエンバクを除いてサポニンの生合成を行うことが出来ないと考えられています (1-4)。双子葉では、トマト、朝鮮人参、甘草など多くの植物に見いだすことができます。サポニンは配糖化トリテルペンのですが、その化学構造や機能から抗菌性サポニンと機能性サポニンに分類されます。私たちはそのうちの抗菌性サポニンに注目し、遺伝子工学的手法を駆使してサポニン非生産性単子葉植物であるイネに抗菌性サポニンの生産能力の賦与を目指します。もちろん食料であるコメへの機能性サポニンの生産能力の賦与も視野に入れています。
すでにイギリスジョンイネスセンターのアン•オズボーン教授らのグループにより、エンバクやトマトでは遺伝子機能解析、LC/MSやGC/MSを用いた抗菌性サポニンの化学分析などが行われています。エンバクやトマトのサポニンの化学的特徴やその生合成経路に関する知見が数多く得られています。(図1、2) (4-8)。
またエンバクやトマト植物などでは植物—病原菌間の認識・感染成立が植物の生産する抗菌性サポニン(アベナシンサポニンやトマチンサポニン)を病原菌が分解できるかどうかにより決定されます (4, 8, 11)。

参考文献

(1) Crombie, L., Crombie, W. M. L. & Whiting, D. A. (1986) J. Chem. Soc. Perkin Trans. 1, 1917–1922.
(2) Hostettmann, K. A., Marston, A. (1995) Saponins. Chemistry and pharmacology of natural products. Cambridge Univ. Press, Cambridge, U.K.
(3) Mary, W., Crombie L., Crombie L. (1986) Distribution of avenacins A-1, A-2, B-1 and B-2 in oat roots: Their fungicidal activity towards ‘take-all’ fungus. Phytochemistry 25, 2069–2073.
(4) Papadopoulou, K., Melton, R. E., Leggett, M. Daniels, M. J. & Osbourn, A. E. (1999) Compromised disease resistance in saponin-deficient plants. Proc. Natl. Acad. Sci. USA 96, 12923–12928.
(5) Qi X, Bakht S, Qin B, Leggett M, Hemmings A, Mellon F, Eagles J, Werck-Reichhart D, Schaller H, Lesot A, Melton R, Osbourn A. (2006) A different function for a member of an ancient and highly conserved cytochrome P450 family: From essential sterols to plant defense. Proc Natl Acad Sci USA 103: 18848-18853.
(6) Haralampidis K, Bryan G, Qi X, Papadopoulou K, Bakht S, Melton R, Osbourn A. (2001) A new class of oxidosqualene cyclases directs synthesis of antimicrobial phytoprotectants in monocots. Proc Natl Acad Sci USA 98:13431-13436.
(7) Qi X, Bakht S, Leggett M, Maxwell C, Melton R, Osbourn A. (2004) A gene cluster for secondary metabolism in oat: implications for the evolution of metabolic diversity in plants. Proc Natl Acad Sci USA 101:8233-8238.
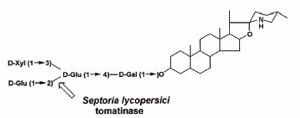
(8) Martin-Hernandez AM, Dufresne M, Hugouvieux V, Melton R, Osbourn A. (2000) Effects of targeted replacement of the tomatinase gene on the interaction of Septoria lycopersici with tomato plants. Mol Plant Microbe Interact. 13:1301-1311.
(9) Briskin DP. (2000) Medical plants and phytomedicines. Linking plant biochemistry and physiology to human health. Plant Physiol. 124: 507-514.
(10) Lee M-H, Jeong J-H, Seo J-W, Shin C-G, Kim Y-S, In J-G, Yang D-C, Yi J-S, Choi Y-E. (2004) Enhanced triterpene and phytosterol biosynthesis in Panax ginseng overexpressing squalene synthase gene. Plant Cell Physiol. 45: 976-984.
(11) Bouarab K, Melton R, Peart J, Baulcombe D, Osbourn A. (2002) A saponin-detoxifying enzyme mediates suppression of plant defences. Nature 418:889-892.
(12) Inagaki Y, Etherington G, Geisler K, Field B, Ikeda K, Mutsukado Y, Dicks J, Osbourn A: (2011) Investigation of the potential for triterpene synthesis in rice through genome mining and metabolic engineering. New Phytologist. 191:432-448.
(13) Inagaki Y, Noutoshi Y, Fujita K, Imaoka A, Arase S, Toyoda K, Shiraishi T, Ichinose Y (2013) Infection-inhibition activity of avenacin saponins against the cereal pathogens Blumeria graminis f.sp. hordei, Bipolaris oryzae, and Magnaporthe oryzae. J. Gen. Plant Pathol. 79(1): 69-73.
(14) 特許「プラントアクティベーター」:特許番号6455833号 (平成30年12月38日登録).


イネにおける抗菌性二次代謝産物:トリテルペン生合成経路遺伝子群の同定と機能解析
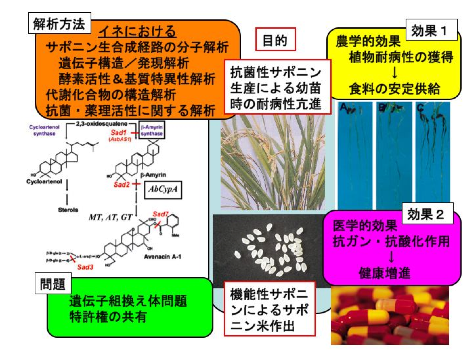
私たちはこのエンバクの知見、イネのゲノム情報等の先端的知見、網羅的遺伝子発現解析方法(マイクロアレイやEST解析)、近年急速に進化しているメタボロミクスによる代謝産物の化学構造分析法なとの最先端技術を活用してイネにおける抗菌性、機能性サポニン生合成経路の解析を試みることによりイネ植物の発芽•幼苗時における病原性糸状菌に対する耐病性の獲得とコメへの機能性サポニン蓄積による新規高機能性米の作出を目指しています(12- 14)。
得られる新規機能としてはイネ育種にとって幼苗時の糸状菌病に対する耐病性獲得に有効に働くことができる可能性が期待されます。もちろん新規機能性食品としてのコメという新しい商品価値(新形質)を賦与することが出来ることも期待できます(図3)。(稲垣)
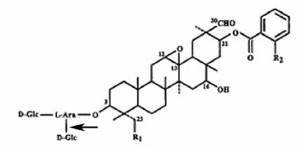
(図1)エンバクが生産する抗菌性アベナシンサポニンの化学構造
(図2)トマトが生産する抗菌性トマチンサポニンの化学構造
(図3)イネにおける抗菌性サポニン生合成に関する分子解析
特に朝鮮人参、甘草、キキョウが生産する機能性サポニンでは抗酸化作用や抗ガン作用など数多くの有用な生理学的機能を有することも知られています (9-10)。機能性サポニンも基本的な化学構造の骨格は配糖化トリテルペンです。
-
 研究にようこそ へ戻る
研究にようこそ へ戻る